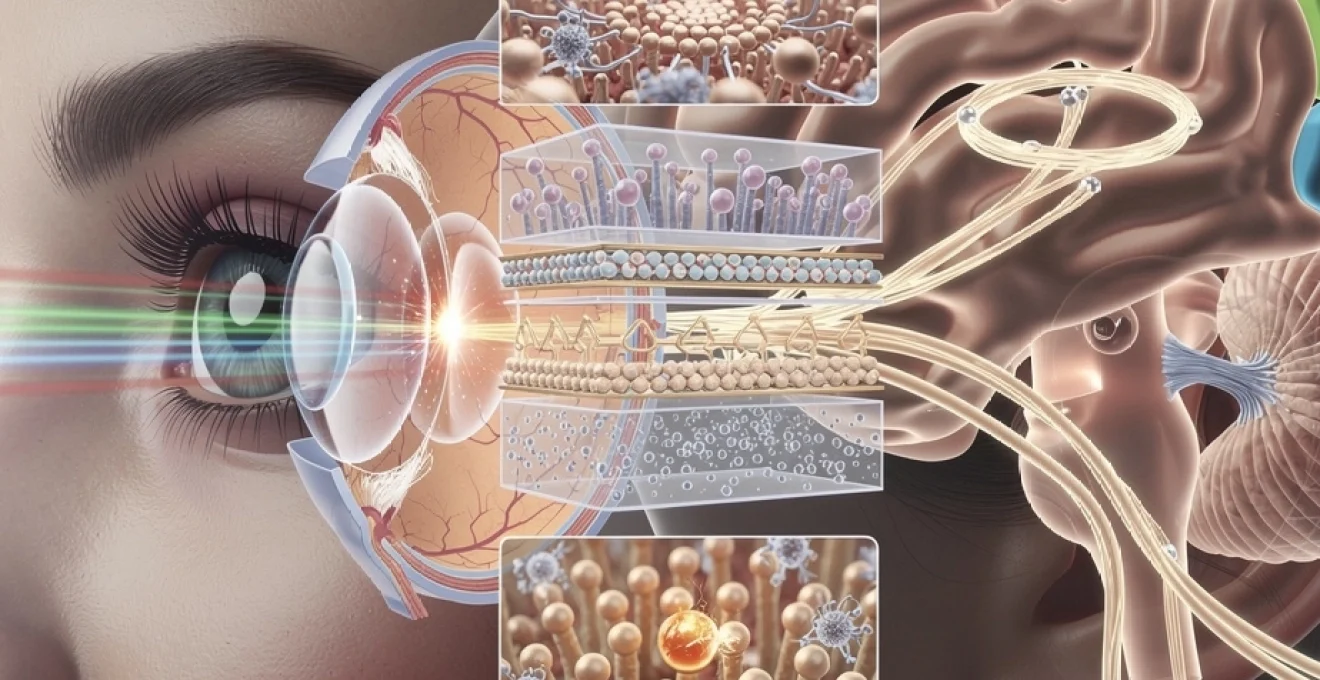
La vision constitue l’un des systèmes sensoriels les plus complexes et fascinants du corps humain. Chaque seconde, vos yeux captent des millions de photons lumineux qui voyagent à travers différentes structures oculaires avant d’être convertis en signaux électriques interprétables par votre cerveau. Ce processus extraordinaire, qui semble instantané et naturel, repose en réalité sur une cascade d’événements biochimiques et neurologiques d’une précision remarquable. La transformation de la lumière en perception visuelle implique non seulement les composants anatomiques de l’œil, mais également des voies nerveuses sophistiquées et des aires cérébrales spécialisées qui travaillent en parfaite synergie. Comprendre ce mécanisme permet d’apprécier la complexité de notre perception quotidienne et d’identifier les dysfonctionnements possibles du système visuel.
L’anatomie structurelle de l’œil : de la cornée à la rétine
L’œil humain peut être comparé à un instrument optique de haute précision, dont chaque composant joue un rôle spécifique dans la formation d’une image nette. La structure anatomique de l’œil s’organise en plusieurs couches et segments, depuis les interfaces externes qui captent la lumière jusqu’aux structures profondes qui la transforment en signaux nerveux. Cette architecture complexe garantit que les photons lumineux suivent un trajet optimal pour produire une image précise sur la rétine.
Le rôle réfractif de la cornée et du cristallin dans la focalisation lumineuse
La cornée représente la première interface que rencontre la lumière en pénétrant dans votre œil. Cette membrane transparente, d’environ 0,5 millimètre d’épaisseur, possède un pouvoir réfractif considérable de 43 dioptries, soit environ deux tiers du pouvoir de réfraction total de l’œil. Sa courbure régulière et sa transparence cristalline sont essentielles pour focaliser correctement les rayons lumineux vers l’intérieur de l’œil. La cornée doit rester constamment hydratée et lisse grâce au film lacrymal qui la recouvre.
Le cristallin, situé juste derrière l’iris, complète le système optique avec une capacité unique : l’accommodation. Cette lentille biconvexe naturelle peut modifier sa courbure grâce aux muscles ciliaires qui l’entourent. Lorsque vous regardez un objet proche, le cristallin se bombe pour augmenter son pouvoir réfractif d’environ 15 dioptries supplémentaires. À l’inverse, pour la vision de loin, il s’aplatit et se détend. Cette flexibilité diminue progressivement avec l’âge, expliquant l’apparition de la presbytie après 40 ans.
La pupille et l’iris : mécanismes de régulation de l’intensité lumineuse
La pupille fonctionne comme un diaphragme photographique automatique, ajustant constamment son diamètre pour réguler la quantité de lumière entrant dans l’œil. Son diamètre peut varier de 2 millimètres en pleine lumière (myosis) à 8 millimètres dans l’obscurité (mydriase), soit une variation d’intensité lumineuse d’un facteur 16. Cette modulation protège les photorécepteurs rétiniens d’une surexposition potentiellement dommageable tout en optimisant la qualité de l’image formée.
L’iris contrôle cette ouverture pupillaire grâce à deux
muscles : un muscle sphincter circulaire qui contracte la pupille en lumière forte, et un muscle dilatateur radial qui l’agrandit dans la pénombre. Ces ajustements se font en quelques fractions de seconde, sous le contrôle du système nerveux autonome. Ce réflexe pupillaire n’agit pas seulement comme un « variateur de lumière », il améliore aussi la profondeur de champ de l’œil, un peu comme lorsque vous fermez légèrement le diaphragme d’un appareil photo pour obtenir une image plus nette.
La synchronisation des deux pupilles est également remarquable : même si vous éclairez un seul œil, la pupille de l’autre œil se contracte aussi, via des circuits réflexes bilatéraux. En pratique, cette capacité d’adaptation à l’intensité lumineuse, combinée à l’accommodation du cristallin, permet à l’œil humain de fonctionner dans des conditions d’éclairement extrêmement diverses, de la nuit faiblement éclairée au plein soleil d’été.
L’humeur aqueuse et le corps vitré : milieux transparents de transmission optique
Entre la cornée et le cristallin se trouve une solution claire, l’humeur aqueuse, produite en continu par les procès ciliaires. Ce liquide nourrit les structures avasculaires de l’avant de l’œil (cornée, cristallin) et participe au maintien de la pression intraoculaire. Sa transparence doit être parfaite pour que la lumière traverse sans être déviée de façon anarchique. Un déséquilibre de sa production ou de son évacuation peut entraîner une élévation de pression, comme dans le glaucome, perturbant à terme la transmission des informations visuelles.
Derrière le cristallin, la majeure partie du globe oculaire est remplie par le corps vitré, un gel transparent composé principalement d’eau, de collagène et d’acide hyaluronique. Il stabilise la forme de l’œil et maintient la rétine appliquée contre la paroi interne. Là encore, la transparence est cruciale : des opacités du vitré, comme les « mouches volantes » (myodésopsies), peuvent projeter des ombres sur la rétine et gêner la vision, sans toutefois bloquer le trajet général de la lumière.
On peut comparer ces milieux transparents à l’intérieur d’un objectif de caméra : s’ils se troublent (cataracte pour le cristallin, hémorragie dans le vitré, œdème cornéen), l’image qui se forme sur le capteur – ici, la rétine – devient floue ou déformée. Préserver leur clarté par une bonne hygiène visuelle, un suivi ophtalmologique régulier et une protection face aux UV est essentiel pour maintenir une bonne qualité d’image rétinienne tout au long de la vie.
La macula et la fovéa : zones de concentration maximale des photorécepteurs
Au fond de l’œil, la lumière se focalise sur une petite zone spécialisée de la rétine : la macula. Cette région jaunâtre d’environ 5 millimètres de diamètre est responsable de la vision fine et de la perception précise des détails. En son centre se trouve la fovéa (ou fovéa centralis), une dépression minuscule de 0,3 millimètre de diamètre où la densité de cônes est maximale et où les bâtonnets sont absents.
La fovéa fonctionne comme le point AF central d’un appareil photo haute résolution : c’est là que vous « posez » votre regard lorsque vous lisez, reconnaissez un visage ou distinguez une petite inscription au loin. La mosaïque de photorécepteurs y est tellement serrée que l’acuité visuelle peut atteindre 10/10, voire davantage chez certains sujets. La moindre altération de cette zone, comme dans la dégénérescence maculaire liée à l’âge (DMLA), entraîne donc une baisse marquée de la vision centrale, alors que la vision périphérique reste souvent préservée.
En périphérie de la macula, la densité de cônes diminue au profit des bâtonnets, mieux adaptés à la vision nocturne et à la détection du mouvement. Ce gradient spatial permet à l’œil humain de combiner une vision centrale extrêmement fine et une vision périphérique large et sensible, indispensable par exemple pour repérer un mouvement sur le côté tout en gardant un texte en lecture au centre du champ visuel.
La phototransduction rétinienne : conversion des photons en signaux électriques
Une fois focalisée sur la rétine, la lumière doit être convertie en signaux électriques pour que le cerveau puisse l’interpréter. Ce processus, appelé phototransduction, transforme l’énergie des photons en variations de potentiel membranaire au niveau des photorécepteurs. Il s’agit d’un véritable « traducteur » : d’un côté, un signal physique (lumière), de l’autre, un langage électrochimique que le système nerveux sait traiter.
Cette conversion ne se fait pas de manière brute. Elle repose sur une architecture cellulaire sophistiquée et sur des cascades biochimiques précises, extrêmement rapides et finement régulées. En quelques millisecondes, une poignée de photons peut ainsi déclencher un signal suffisamment fort pour être relayé puis traité par l’ensemble du système visuel. C’est cette efficacité qui explique que nous puissions distinguer une étoile dans un ciel nocturne ou percevoir des nuances subtiles au sein d’une scène très éclairée.
Les cônes et bâtonnets : architecture moléculaire des photorécepteurs
Les photorécepteurs de la rétine se répartissent en deux grandes catégories : les bâtonnets et les cônes. On dénombre environ 120 millions de bâtonnets et 6 à 7 millions de cônes dans un œil humain. Les bâtonnets, très sensibles à la faible luminosité, dominent la périphérie rétinienne et assurent la vision scotopique (vision nocturne) ainsi que la perception des mouvements. Les cônes, concentrés dans la macula et surtout dans la fovéa, sont responsables de la vision des couleurs et de l’acuité visuelle élevée.
Chaque photorécepteur possède trois grandes parties : un segment externe où se trouvent des disques membranaires riches en photopigments, un segment interne contenant les organites nécessaires au métabolisme cellulaire, et une terminaison synaptique qui communique avec les cellules bipolaires. Dans les bâtonnets, le photopigment principal est la rhodopsine, tandis que les cônes renferment des opsines spécifiques (S, M, L) sensibles à différentes longueurs d’onde.
On peut comparer ces photorécepteurs à des capteurs d’un appareil photo numérique, mais avec une sophistication supplémentaire : chaque type de « capteur » possède un profil spectral distinct et une sensibilité variable, ce qui permet non seulement de détecter la quantité de lumière, mais aussi de coder sa composition spectrale. Sans cette architecture moléculaire fine, la lumière resterait un simple flux énergétique et ne pourrait être traduite en informations visuelles détaillées.
Le cycle de la rhodopsine et des opsines : mécanisme biochimique d’absorption photonique
Au cœur de la phototransduction se trouve l’absorption d’un photon par un photopigment. La rhodopsine, dans les bâtonnets, et les différentes opsines, dans les cônes, sont des protéines transmembranaires associées à un chromophore, le 11-cis-rétinal, dérivé de la vitamine A. Lorsque ce chromophore absorbe un photon, il change instantanément de conformation pour devenir du tout-trans-rétinal. Ce simple basculement moléculaire agit comme un interrupteur qui déclenche toute la cascade de transduction.
Cette forme activée de la rhodopsine (ou des opsines) est souvent appelée métarhodopsine II. Elle interagit alors avec une protéine G spécifique, la transducine, initiant une amplification considérable du signal initial. Quelques photons suffisent à activer de nombreuses molécules de transducine, ce qui fait des photorécepteurs des détecteurs extrêmement sensibles. Par la suite, le rétinal est dissocié de l’opsine et doit être recyclé via le cycle visuel impliquant l’épithélium pigmentaire rétinien.
Ce cycle de régénération des photopigments est indispensable pour maintenir une vision continue. En cas de déficit grave en vitamine A ou de dysfonctionnement de l’épithélium pigmentaire, la régénération est compromise, entraînant des troubles visuels, en particulier en vision nocturne. On comprend alors mieux pourquoi une alimentation équilibrée et la santé générale de la rétine sont si importantes pour la transformation correcte de la lumière en signal nerveux.
La cascade de transduction par la protéine G et la phosphodiestérase
Une fois activée, la rhodopsine stimule la transducine, qui à son tour active une enzyme clé : la phosphodiestérase (PDE). Cette enzyme hydrolyse le GMPc (guanosine monophosphate cyclique), un second messager qui, dans l’obscurité, maintient ouverts des canaux ioniques sensibles au GMPc dans la membrane du segment externe. Tant que le GMPc est abondant, les canaux sodiques et calciques restent ouverts, permettant un flux constant d’ions dans la cellule.
Lorsque la PDE réduit rapidement la concentration de GMPc, ces canaux se ferment. La cellule photoréceptrice passe alors d’un état relativement dépolarisé (dans le noir) à un état plus polarisé (en présence de lumière). Ce changement de potentiel membranaire modifie la libération de neurotransmetteurs au niveau de la terminaison synaptique, signalant ainsi la présence de lumière aux cellules bipolaires. On peut voir cette cascade comme un système d’amplification : un photon entraîne l’activation de nombreuses molécules enzymatiques et la fermeture d’un grand nombre de canaux ioniques.
La précision de cette cascade de transduction est telle que l’œil peut distinguer des variations d’éclairement sur plusieurs ordres de grandeur. Des mécanismes de régulation, impliquant notamment le calcium intracellulaire, permettent d’ajuster en permanence la sensibilité des photorécepteurs. Sans ces boucles de rétrocontrôle, nous serions éblouis en permanence en passant d’un environnement sombre à un environnement lumineux.
L’hyperpolarisation membranaire : inversion du signal neuronal
Contrairement à la plupart des neurones, qui se dépolarisent lorsqu’ils sont activés, les photorécepteurs s’hyperpolarisent en réponse à la lumière. Dans l’obscurité, ils libèrent en continu un neurotransmetteur, le glutamate, au niveau de leurs synapses avec les cellules bipolaires. Lorsque la lumière provoque la fermeture des canaux ioniques et l’hyperpolarisation, cette libération de glutamate diminue proportionnellement à l’intensité lumineuse.
Cette « inversion » du signal neuronal peut paraître contre-intuitive, mais elle représente une stratégie très efficace pour coder les variations de lumière plutôt que la lumière absolue. Le système visuel est en effet particulièrement sensible aux contrastes et aux changements, plus qu’aux niveaux d’éclairement statiques. En pratique, cela permet à l’œil d’économiser de l’énergie et de maximiser sa réactivité aux transitions lumineuses rapides.
Ce signal modulé est ensuite transmis aux cellules bipolaires, qui interprètent différemment la baisse de glutamate selon leur type (ON ou OFF). Déjà à ce stade, l’information lumineuse commence à être organisée en motifs de luminosité croissante ou décroissante, préparant ainsi le traitement plus complexe qui sera réalisé dans les couches rétiniennes suivantes et dans le cortex visuel.
Le traitement neural dans les couches rétiniennes
La rétine n’est pas un simple « film passif » recevant l’image. C’est un véritable mini-cerveau qui effectue un premier niveau de traitement de l’information visuelle. Avant même que les signaux n’atteignent le cerveau, la rétine filtre, compresse et met en forme les données lumineuses. Elle détecte notamment les contrastes, les contours, certains mouvements et des variations locales de luminosité.
Cette organisation repose sur plusieurs couches cellulaires superposées : les photorécepteurs, les cellules bipolaires, horizontales, amacrines et ganglionnaires. À travers un réseau sophistiqué de connexions excitatrices et inhibitrices, la rétine construit des modèles d’activité qui correspondent déjà à des unités d’information visuelle utiles, comme les « points lumineux sur fond sombre » ou inversement. C’est grâce à ce pré-traitement que le nerf optique peut transmettre une information déjà partiellement interprétée, plutôt qu’un simple flux brut de données.
Les cellules bipolaires : premier relais de transmission synaptique
Situées entre les photorécepteurs et les cellules ganglionnaires, les cellules bipolaires constituent le premier relais synaptique de la voie visuelle. Elles reçoivent le signal modulé par la lumière (variation de glutamate) et le traduisent en potentiels gradués. Il existe plusieurs types de cellules bipolaires, notamment les bipolaires ON et OFF, qui répondent de manière opposée à une augmentation de lumière.
Les cellules bipolaires ON sont activées lorsque la lumière augmente, tandis que les cellules OFF sont stimulées lorsque la lumière diminue. Ce découpage du signal en deux voies complémentaires permet un codage plus précis des changements locaux de luminance. En termes d’analogie, on peut considérer que la rétine crée dès ce stade deux « canaux » distincts : l’un spécialisé dans la détection des points plus clairs que leur environnement, l’autre dans la détection des points plus sombres.
Les cellules bipolaires collectent également les signaux de plusieurs photorécepteurs, surtout en vision périphérique, ce qui augmente la sensibilité mais réduit la précision spatiale. À l’inverse, au niveau de la fovéa, la convergence est minimale : un cône peut se connecter à une seule cellule bipolaire, puis à une cellule ganglionnaire unique, garantissant une résolution spatiale maximale, cruciale pour la lecture ou la reconnaissance de visages.
Les cellules ganglionnaires et leurs champs récepteurs
Les cellules ganglionnaires constituent la dernière couche neuronale de la rétine et leurs axones forment le nerf optique. Elles transforment les signaux gradués des cellules bipolaires en potentiels d’action, c’est-à-dire en impulsions électriques codées en fréquence, adaptées au transport sur de longues distances jusqu’au cerveau. Chaque cellule ganglionnaire répond à un motif spatial particulier dans une petite zone du champ visuel appelée champ récepteur.
La plupart des champs récepteurs des cellules ganglionnaires sont organisés de façon centre-périphérie (ou centre-surround). Par exemple, une cellule de type ON-centre / OFF-périphérie sera fortement activée par un point lumineux au centre entouré d’une zone plus sombre, et inhibée si la lumière éclaire principalement la périphérie. Cette organisation permet une détection très fine des contrastes locaux et des contours d’objets.
Il existe aussi différentes classes fonctionnelles de cellules ganglionnaires, comme les cellules parvocellulaires (spécialisées dans les détails et les couleurs) et magnocellulaires (sensibles au mouvement et aux variations rapides de luminance). Ainsi, dès la sortie de la rétine, l’information visuelle est déjà séparée en canaux parallèles qui seront traités différemment par les structures cérébrales ultérieures.
Les cellules horizontales et amacrines : modulation latérale du signal
En parallèle des voies directes photorécepteur → bipolaire → ganglionnaire, la rétine possède des circuits latéraux assurés par les cellules horizontales et les cellules amacrines. Les cellules horizontales, situées entre les photorécepteurs et les bipolaires, intègrent des informations sur une zone plus large et exercent une influence inhibitrice latérale. Ce mécanisme de lateral inhibition accentue les contrastes aux frontières entre zones claires et sombres, renforçant la perception des bords.
Les cellules amacrines, interposées entre les cellules bipolaires et ganglionnaires, forment une famille très diverse de neurones internes. Certaines sont spécialisées dans la détection de mouvements directionnels, d’autres régulent la sensibilité globale de la rétine ou participent à l’adaptation à la lumière. On peut voir ces cellules comme un réseau de « réglages fins » qui optimise en permanence la qualité de l’information envoyée au cerveau.
Ces mécanismes de modulation latérale expliquent de nombreux phénomènes visuels du quotidien, comme les illusions d’optique liées aux contrastes exagérés ou à la perception de contours illusoires. Sans ce pré-traitement sophistiqué, notre vision serait beaucoup plus plate, moins sensible aux contours et aux variations de lumière qui structurent notre environnement visuel.
Les voies optiques cérébrales : du nerf optique au cortex visuel
Une fois codée par les cellules ganglionnaires, l’information visuelle quitte l’œil par le nerf optique. Chaque œil envoie environ un million de fibres nerveuses vers le cerveau, formant ainsi un câble de données à très haut débit. Ce signal ne se contente pas de « monter » directement vers le cortex : il transite par plusieurs relais anatomiques qui filtrent, organisent et synchronisent les informations issues des deux yeux.
Ce trajet, qui va de la rétine jusqu’au cortex occipital, est appelé voie visuelle. Il comprend notamment le chiasma optique, les bandelettes optiques, le corps genouillé latéral du thalamus et les radiations optiques. À chaque étape, une partie du traitement est déjà réalisée, préparant le cortex visuel primaire à recevoir une carte du monde extérieur aussi fidèle et organisée que possible.
Le chiasma optique : décussation partielle des fibres nerveuses
Les deux nerfs optiques se rejoignent au niveau du chiasma optique, structure située à la base du cerveau, juste en avant de l’hypophyse. À cet endroit, une partie des fibres nerveuses croise la ligne médiane. Plus précisément, les fibres issues de la moitié nasale de chaque rétine décussent, tandis que celles issues de la moitié temporale restent du même côté.
Pourquoi ce croisement partiel est-il si important ? Il permet à chaque hémisphère cérébral de recevoir les informations visuelles en provenance du champ visuel contralatéral (gauche ou droit). Ainsi, l’hémisphère droit traite principalement ce que vous voyez dans votre champ visuel gauche, et inversement. Cette organisation est essentielle pour la perception binoculaire et la vision en trois dimensions.
Des lésions au niveau du chiasma optique, par exemple dues à une tumeur hypophysaire comprimant cette structure, provoquent des défauts du champ visuel caractéristiques (comme une hémianopsie bitemporale). Comprendre cette décussation partielle permet d’interpréter les symptômes visuels et de localiser les atteintes neurologiques avec précision.
Le corps genouillé latéral du thalamus : relais sous-cortical majeur
Après le chiasma optique, les fibres continuent leur trajet dans les bandelettes optiques jusqu’au corps genouillé latéral (CGL), un noyau du thalamus spécialisé dans la vision. Le CGL agit comme une « station de tri » et de modulation : il reçoit les signaux rétiniens, les organise en couches distinctes et les ajuste en fonction de l’état général de l’organisme (attention, veille, sommeil).
Le CGL est composé de six couches principales chez l’humain : deux couches magnocellulaires (sensibles au mouvement et aux variations rapides) et quatre couches parvocellulaires (dédiées aux détails fins et aux couleurs). Les informations provenant de chaque œil restent séparées au sein de ces couches, ce qui permet de conserver une représentation précise de la dominance oculaire et de préparer la fusion binoculaire dans le cortex.
On peut comparer le CGL à un hub de routage intelligent qui, en plus de transmettre l’information, la filtre et la pondère. Par exemple, en situation de forte attention visuelle, certains signaux peuvent être amplifiés, tandis que d’autres, jugés moins pertinents, sont atténués. Ce contrôle thalamique illustre à quel point la vision n’est pas seulement un phénomène passif, mais un processus actif, influencé par l’état mental et les besoins cognitifs.
Les radiations optiques et leur projection vers le cortex occipital
Depuis le corps genouillé latéral, les signaux visuels sont envoyés vers le cortex occipital via un large faisceau de fibres appelé radiations optiques. Ces fibres se déploient en éventail à travers le lobe temporal et le lobe pariétal avant de rejoindre la face médiale du lobe occipital, où se trouve le cortex visuel primaire (aire 17 de Brodmann).
Les radiations optiques respectent une organisation topographique stricte : les différentes régions du champ visuel sont représentées dans des portions spécifiques de ce faisceau. Par exemple, les informations provenant du quadrant visuel supérieur transitent par une boucle plus inférieure (boucle de Meyer) dans le lobe temporal. Des lésions focales sur ce trajet entraînent des déficits du champ visuel caractéristiques (quadranopsies), ce qui aide les neurologues à localiser précisément les atteintes cérébrales.
Une fois arrivées dans le cortex occipital, ces projections conservent l’organisation spatiale héritée de la rétine, permettant au cerveau de construire une « carte » du monde extérieur. C’est la base de la représentation rétinotopique, sur laquelle vont se greffer des traitements de plus en plus complexes pour reconnaître les formes, les couleurs et le mouvement.
Le cortex visuel primaire V1 : cartographie rétinotopique et colonnes de dominance oculaire
Le cortex visuel primaire (V1) est la première région corticale à recevoir directement les signaux des radiations optiques. Il est organisé en une carte rétinotopique où chaque point du champ visuel correspond à une zone précise du cortex. La fovéa, très riche en photorécepteurs, occupe une surface disproportionnée dans V1, reflétant l’importance de la vision centrale pour les tâches fines.
Au sein de V1, les neurones sont regroupés en colonnes de dominance oculaire : des micro-unités verticales qui répondent préférentiellement aux signaux provenant de l’un ou l’autre œil. Cette organisation permet la comparaison entre les deux entrées oculaires, essentielle à la stéréoscopie (perception du relief). V1 est également structuré en colonnes d’orientation, chaque colonne répondant à des bords ou des lignes ayant une orientation particulière (verticale, horizontale, oblique).
On peut voir V1 comme un réseau de détecteurs de « primitives visuelles » : contrastes, orientations, positions dans le champ visuel. À ce stade, le cerveau n’a pas encore « reconnu » des objets au sens où nous l’entendons, mais il a déjà décomposé la scène en éléments géométriques simples qui serviront de briques de base pour les aires visuelles supérieures.
Le traitement cortical supérieur : perception et intégration visuelle
À partir de V1, l’information visuelle est transmise à un ensemble d’aires dites extrastriées (V2, V3, V4, V5/MT, etc.) qui réalisent des traitements de plus en plus élaborés. C’est dans ces régions que la mosaïque de lignes, de contrastes et de mouvements issue de V1 est progressivement intégrée en formes, objets, visages et scènes cohérentes.
Ce traitement n’est pas linéaire : il s’effectue en parallèle dans plusieurs voies spécialisées et implique de nombreuses boucles de rétroaction. Le cerveau compare en permanence les informations entrantes avec vos souvenirs, vos attentes et votre contexte émotionnel. C’est pourquoi deux personnes peuvent percevoir différemment une même scène : la perception visuelle est autant une construction cognitive qu’un simple enregistrement de la réalité.
Les aires visuelles extrastriées V2, V3, V4 et V5/MT : spécialisation fonctionnelle
Chaque aire visuelle extrastriée joue un rôle spécifique dans l’analyse de la scène visuelle. L’aire V2 reçoit directement des entrées de V1 et participe à l’intégration des contours et des textures. L’aire V3 est impliquée dans le traitement de la structure globale des formes et de certaines informations de profondeur. L’aire V4 est particulièrement importante pour la perception des couleurs et des formes complexes.
L’aire V5 ou MT (middle temporal) est, quant à elle, spécialisée dans la détection du mouvement. Les neurones de V5 sont sensibles à la direction et à la vitesse des objets en déplacement. C’est notamment grâce à cette aire que vous pouvez suivre une balle en mouvement ou percevoir la direction d’un véhicule sur la route. Des lésions focales de V5 peuvent entraîner une acinetopsie, trouble rare dans lequel la perception du mouvement est gravement altérée.
On peut comparer ces aires extrastriées à des « services spécialisés » au sein d’une entreprise : chacune traite un type d’information (couleur, forme, mouvement) mais toutes collaborent pour produire une perception unifiée. Cette spécialisation fonctionnelle explique aussi pourquoi certains troubles neurologiques touchent sélectivement la reconnaissance des visages, des lieux ou des mouvements, tout en laissant relativement intacte la vision globale.
La voie dorsale « où » et la voie ventrale « quoi » : modèle de ungerleider et mishkin
Les recherches en neurosciences ont montré que les informations visuelles se répartissent principalement en deux grandes voies de traitement, décrites par Ungerleider et Mishkin : la voie dorsale et la voie ventrale. La voie dorsale, qui part de V1 et V2 pour remonter vers les régions pariétales, est souvent qualifiée de voie du « où » ou du « comment ». Elle est impliquée dans la localisation spatiale des objets, la perception du mouvement et la coordination visuo-motrice.
La voie ventrale, quant à elle, descend de V1 et V2 vers les régions temporales inférieures et est appelée voie du « quoi ». Elle est spécialisée dans la reconnaissance des formes, des objets, des visages et des scènes. C’est grâce à cette voie que vous pouvez reconnaître instantanément une chaise, un visage familier ou la façade de votre maison. Les deux voies fonctionnent de concert : pour saisir un objet, vous devez à la fois savoir ce que c’est et où il se trouve dans l’espace.
Des lésions sélectives de ces voies illustrent leur rôle spécifique. Une atteinte de la voie ventrale peut entraîner une agnosie visuelle (incapacité à reconnaître des objets malgré une acuité conservée), tandis qu’une atteinte de la voie dorsale peut provoquer des difficultés à guider les gestes par la vision. Cette distinction souligne encore une fois que transformer la lumière en perception visuelle compréhensible ne se limite pas à « voir net » : il s’agit d’interpréter, de localiser et d’agir en fonction de ce que l’on voit.
Le traitement des contours, orientations et fréquences spatiales
Un aspect fondamental du traitement cortical supérieur concerne l’analyse des contours et des fréquences spatiales. Les neurones de V1 et des aires voisines sont sensibles à des motifs précis : lignes d’une certaine orientation, bords à contraste élevé, motifs répétés à différentes échelles. On peut comparer cette analyse à un ensemble de filtres qui décomposent l’image en composantes de basse fréquence (formes globales) et de haute fréquence (détails fins).
Les basses fréquences spatiales correspondent aux variations lentes de luminosité et de couleur, utiles pour percevoir la structure générale d’une scène (par exemple, distinguer le ciel du sol). Les hautes fréquences, au contraire, correspondent aux détails comme les feuilles d’un arbre ou les traits d’un visage. Le cerveau combine ces informations pour construire une perception à la fois globale et détaillée. C’est ce qui vous permet, par exemple, de reconnaître quelqu’un de loin grâce à sa silhouette (basses fréquences), puis d’identifier son expression lorsque vous vous approchez (hautes fréquences).
Le traitement des orientations et des fréquences spatiales est également à l’origine de nombreuses illusions visuelles. Certains motifs, comme les grilles à contraste élevé ou les lignes obliques proches, peuvent tromper nos systèmes de détection de contours, créant des perceptions d’ondulation ou de déformation inexistantes. Ces illusions ne sont pas de simples curiosités : elles révèlent le fonctionnement intime des mécanismes cérébraux qui transforment la lumière en formes signifiantes.
La vision colorée trichromatique et l’encodage chromatique
Au-delà de la forme et du mouvement, la couleur joue un rôle majeur dans notre perception visuelle. Elle nous aide à segmenter les objets, à reconnaître des éléments familiers et même à susciter des émotions. Pourtant, comme nous l’avons vu, les objets n’ont pas de couleur en eux-mêmes : ils absorbent et reflètent certaines longueurs d’onde de la lumière, que notre système visuel interprète ensuite comme des couleurs spécifiques.
La vision colorée repose sur la présence de trois types de cônes, sur des circuits rétiniens spécialisés et sur des mécanismes cortico-thalamiques d’intégration. Ensemble, ils permettent à l’être humain de distinguer jusqu’à plusieurs millions de nuances différentes. Comment cette richesse chromatique est-elle possible avec seulement trois types de récepteurs ? C’est ce que nous allons voir en détaillant les principales théories et mécanismes d’encodage.
La théorie trichromatique de Young-Helmholtz : trois types de cônes S, M et L
La théorie trichromatique, proposée par Young puis quantitative par Helmholtz, postule que la vision des couleurs repose sur trois types de photorécepteurs coniques, chacun sensible à une gamme différente de longueurs d’onde. Chez l’humain, on distingue les cônes S (Short), plus sensibles au bleu-violet, les cônes M (Medium), sensibles au vert, et les cônes L (Long), sensibles au jaune-rouge.
Chaque cône ne répond pas à une seule longueur d’onde précise, mais à un spectre relativement large, avec un maximum de sensibilité (environ 430 nm pour S, 535 nm pour M et 565 nm pour L). La couleur perçue résulte alors de la combinaison relative des réponses de ces trois types de cônes. Par exemple, une lumière qui active fortement les cônes L et M, mais peu les cônes S, sera perçue comme jaune. C’est le principe même de la synthèse additive des couleurs utilisé dans les écrans et l’éclairage LED.
Cette théorie explique aussi les anomalies trichromatiques comme le daltonisme rouge-vert, où un type de cône (M ou L) manque ou fonctionne anormalement. Dans ces cas, l’espace des couleurs perceptibles est réduit, certaines teintes étant confondues. Des recherches en thérapie génique explorent aujourd’hui la possibilité de restaurer ou d’améliorer cette vision trichromatique chez certains patients, en réintroduisant des opsines manquantes dans les cônes existants.
La théorie des processus opposants de hering : codage antagoniste rouge-vert et bleu-jaune
La théorie trichromatique ne suffit toutefois pas à expliquer certains phénomènes, comme l’impossibilité de percevoir un « rouge-vert » ou un « jaune-bleu » simultané, ou encore la nature particulière des couleurs complémentaires. Pour répondre à ces observations, Hering a proposé la théorie des processus opposants. Selon cette approche, le système visuel ne code pas seulement les réponses des trois types de cônes, mais aussi des différences entre ces réponses dans des canaux antagonistes.
On distingue principalement trois canaux : un canal rouge-vert (opposition entre les signaux des cônes L et M), un canal bleu-jaune (opposition entre les cônes S et la combinaison L+M) et un canal noir-blanc (lié à la luminance globale). Ainsi, lorsqu’un canal rouge-vert est fortement activé dans le sens « rouge », il est simultanément inhibé dans le sens « vert », ce qui rend impossible la perception d’un mélange rouge-vert pur. Ces mécanismes d’opposition apparaissent déjà au niveau des cellules bipolaires et ganglionnaires, puis se prolongent dans les relais thalamiques et corticaux.
Cette organisation en processus opposants explique nombreuses illusions chromatiques, comme les images rémanentes complémentaires (voir du vert après avoir fixé longtemps une surface rouge). Elle montre aussi que la couleur, telle que nous la percevons, est le résultat d’un calcul complexe sur les signaux des cônes, et non un simple reflet des longueurs d’onde présentes dans la lumière.
Les mécanismes d’adaptation chromatique et la constance des couleurs
Vous êtes-vous déjà demandé pourquoi une feuille de papier blanche vous semble blanche aussi bien sous une lumière du jour légèrement bleutée que sous une ampoule à incandescence plutôt jaunâtre ? Ce phénomène, appelé constance des couleurs, repose sur des mécanismes d’adaptation chromatique à plusieurs niveaux du système visuel.
Les photorécepteurs s’adaptent d’abord localement : en présence prolongée d’une teinte dominante, certains cônes réduisent leur sensibilité relative, rééquilibrant ainsi la perception. Ensuite, les circuits rétiniens et thalamiques ajustent les réponses des canaux opposants en tenant compte de la distribution globale des couleurs dans la scène. Enfin, les aires corticales supérieures intègrent les connaissances et les attentes : vous « savez » qu’une feuille est blanche, ce qui influence inconsciemment votre interprétation des signaux lumineux.
Cette capacité d’adaptation est cruciale pour que la perception reste stable dans des environnements d’éclairage très variés. Sans elle, chaque changement de source lumineuse transformerait radicalement les couleurs perçues, rendant le monde difficilement reconnaissable. Elle a également des implications pratiques : en photographie, en design ou en ergonomie des écrans, il est essentiel de tenir compte de la façon dont l’œil humain s’adapte à la lumière pour proposer des environnements visuels confortables et fidèles à la réalité.